|
|||||||||||||||||
|
|||||||||||||||||
| Género Amblyopone Erichson, 1842
©Kiko Gómez & Xavier Espadaler |
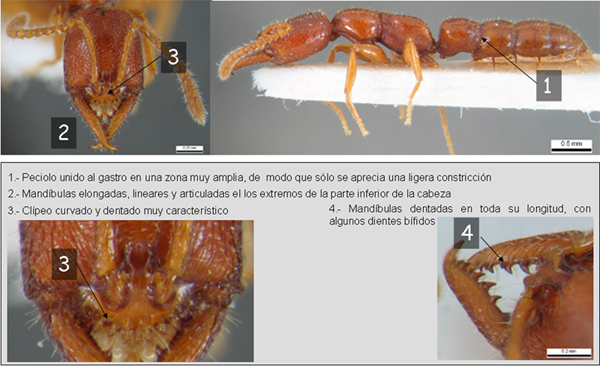
Género inconfundible en la Península Ibérica debido a la forma del peciolo y las mandíbulas características.
Tinaut (1988a) realiza una revisión de las citas de este género en la Península, encontrando que las especies presentes son A. emeryi y A. gaetulica. Otras especies citadas para la Península como A. denticulata (Collingwood & Yarrow 1969) han resultado ser identificaciones falsas.
Género de distribución mundial excepto en Madagascar, sur de la región Etiópica y norte de la Paleártica. Se conocen mas de 60 especies en todo el mundo. En la Península Ibérica se ha localizado en el suroeste peninsular, sin sobrepasar los 39ºN, 4ºW.
Este es un género bastante primitivo, que presenta características poco evolucionadas dentro de las hormigas. Son especies fundamentalmente hipogeas que viven en colonias pequeñas, bajo rocas o en la madera en descomposición. el tamaño de las colonias es pequeño. De 37 colonias de A. pallipes censadas en Norteamérica, la media de adultos por nido resultó de 12, variando entre 1 y 35 (Francoeur, 1965; Traniello, 1982), mientras que tres nidos de A. pluto en Costa de Marfil arrojaron una media de 33 adultos (Gotwald & Lévieux 1972). A pesar del primitivismo de este género, los nidos suelen dispersarse en primavera y verano para reunirse de nuevo en otoño e hibernar (Herbers 1985). En lo relativo a la fundación de nidos, las reinas todavía salen a forrajear fuera del nido, en lo que se denomina "fundación de colonias parcialmente enclaustradas" (partially claustral colony founding)(Holldobler & Wilson 1990). En las hormigas más evolucionadas, esto no sucede, sino que la reina se queda enclaustrada en el nido y alimenta a las crías con huevos tróficos, esto es, destina parte de la producción de huevos a criar a las larvas, que una vez desarrolladas saldrán a forrajear. En cuanto al número de reinas, existen especies con una sola reina por nido (monogínicas) y especies con varias reinas por nido (poligínicas). Los nidos suelen carecer de una cámara central, estando dispersas en varias cámaras con varias obreras y parte de la prole en cada una de ellas (Gotwald & Lévieux 1972). Incluso en estas hormigas tan antiguas se dan dos comportamientos típicos, como son la alarma (Traniello, 1982) y el cuidado de la prole tanto a los huevos como a larvas y pupas (Holldobler & Wilson 1990). No existe división del trabajo por castas, más evolucionado, sino que se da el "polietismo temporal" o división del trabajo en función de la edad. Existe una especie, de hecho (A. pallipes) que ni siquiera presenta esta división (Traniello, 1978; 1982). Asimismo, esta especie en particular son más espontáneos en sus actividades y menos dependientes de la estimulación táctil por parte de otras hormigas que en las colonias de hormigas más evolucionadas, donde se producen explosiones de actividad sincronizada. Cuando una hormiga perteneciente a las especies A. australis o A. pallipes se encuentra con otra de su nido, mueven la cabeza y el torax vigorosamente arriba y abajo. Parece ser que las feromonas segregadas por la glándula mandibular de A. pallipes juegan cierto papel o en el reclutamiento o en las señales de alarma, si bien faltan experimentos que lo concreten (Holldobler, 1977; Traniello, 1982). En esta hormiga también se ha demostrado que emiten feromonas olorosas para guiarse individualmente, si bien no actúan como feromonas de reclutamiento masivo (Holldobler & Wilson 1990). El primitivismo de este género se vuelve a poner de manifiesto en el apareamiento. La hembra, áptera y algunas veces no más que una obrera fértil, permanece en las cercanías del nido y bien en el suelo, bien sobre vegetación baja, emite feromonas que atraerán al macho (Haskins, 1928). Se alimentan de artrópodos que cazan forrajenado en el suelo y no aceptan fluidos azucarados si le son ofrecidos. La mayoría de las especies estudiadas están especializadas en cazar ciempiés como A. pallipes en Norteamérica (Haskins, 1928; Wheeler, 1936; Traniello, 1982) o A. mutica, A. pluto y A. stigia en África Occidental (Gotwald & Lévieux 1972, Lévieux 1983), si bien al menos una especie australiana A. australis caza todo tipo de artrópodos (Haskins & Haskins, 1951). Por norma general, las presas son aguijoneadas paralizándolas y posteriormente son transportadas al nido y almacenadas. En caso de ser molestadas, estas hormigas se mueven lentamente o bien permanecen inmóviles. Existe un tipo de comportamiento extraordinario en estas hormigas, descubierto por Masuko (1986) para A. silvestrii. Las reinas son vampiras que se alimentan de la hemolinfa (la "sangre") de las larvas. Para ello muerden a las larvas más viejas en la parte delantera dorsal del tegumento, y sorben la hemolinfa. Estas reinas no se alimentan ni de huevos tróficos, ni de ciempiés, ni se les regurgita comida por parte de las obreras. Aparte de la herida recibida, las larvas no parecen verse afectadas por este comportamiento.
Las hormigas mediterráneas del género han sido revisadas por Baroni Urbani (1978), y es el texto de base su identificación. En la Península Ibérica es impresindible el trabajo de Tinaut (1990). En otras regiones geográficas existen trabajos recientes como el de Zheng (2001) con claves a las especies del Sureste Asiático, o Lattke (1991) con claves para la fauna del continente americano complementado por Lacau & Delabie (2002) donde se incluyen claves actualizadas del género en la región Neotropical. La Melanesia se revisa en Taylor (1979), proponiendo claves para la identificación de las obreras de la zona. La fauna africana de este género carece de revisiones recientes publicadas, aunque existen claves actualizadas en internet de la región sub-sahariana en Ants of West Africa. Baroni Urbani, C.; 1978; Contributo alla conoscenza del genere Amblyopone; Mitteilungen der Schweizerischen Entomologischen gessellschaft; 51 (1): 39-51 Brown, W. L., Jr.; 1949, A new American Amblyopone, with notes on the genus (Hymenoptera: Formicidae); Psyche; 56: 81-88 Brown, W. L Jr.; 1960; Contributions towards a reclassification of the Formicidae. 3. Tribe Amblyopponini; Bulletin of the Museum of comparative Zoology at Harvard College; 122 (4): 145-230 Francoeur, A.; 1965; Ecologie des populations de fourmis dans un bois de chênes rouges et dérables rouges; Naturaliste Canadien; 92 (10-11): 263-276 Gotwald, W. H., Jr.; Lévieux, J. (1972). Taxonomy and biology of a new West African ant belonging to the genus Amblyopone. Annals of the Entomological Society of America, 65 (2): 383-396 Haskins, C. P.; 1928; Notes on the behaviour and habits of Stigmatomma pallipes Haldeman; Journal of the New York Entomological Society; 36 (2): 179-184 Haskins, C. P.; 1970; Researches in the biology and social behaviour of primitive ants; L. R. Aronson, D. S. Lehrman, and J. S. Rosenblatt eds.; Development and evolution of behaviour; 355-388 Haskins, C. P.; Haskins, E. F.; 1951; Notes on the method of colony foundation of the ponerine ant Amblyopone australis Erichson; American Midland Naturalist; 45 (2): 432-445 Herbers, J. M. (1985). Seasonal structuring of a north temperate ant community. Insectes Sociaux, 32: 224-240 Holldobler, B.; 1977; Comunication in social Hymenoptera; T. A. Sebeok eds.; How animals communicate. Indiana University Press, Bloomington: 418-471 Holldobler, B; Wilson, E. O.; 1990;The Ants; Harvard University Press: 732 pp Lacau, S. ; Delabie, J. H. C. (2002). Description de trois nouvelles espèces d'Amblyopone avec quelques notes biogéographiques sur le genre au Brésil. Bulletin de la Société Entomologique de France, 107 (1): 33-41 Lattke, J. E. (1991). Studies of neotropical Amblyopone Erichson (Hymenoptera: Formicidae).. Contributions in Science, 428: 1-7 Lévieux, J.; 1983;The soil fauna of tropical savannas, IV: The Ants; F. Bourlière eds.; Tropical savannas; 525-540 Masuko, K.; 1986; Larval hemolymph feeding: a nondestructive parental cannibalism in the primitive ant Amblyopone silvestrii Wheeler; Behavioral Ecology and Sociobiology; 19 (4): 249-255 Shattuck, S. O.; 1999;Australian ants: Their biology and identification; Monographs on Invertebrate Taxonomy; Vol 3: 226 p Tinaut, A.; 1988a; El género Amblyopone Erichson en la Península Ibérica; Miscellania Zoologica; 12: 189-193 Traniello, J. F. A.; 1978; Caste in a primitive ant: absence of age polyethism in Amblyopone; Science; 202: 770-772 Traniello, J. F. A.; 1982; Population structure and social organization in the primitive ant Amblyopone pallipes; Psyche; 89(1-2): 65-80 Taylor, R. W. (1979 ("1978") ). Melanesian ants of the genus Amblyopone (Hymenoptera: Formicidae).. Australian Journal of Zoology, 26: 823-839 Wheeler, W. M.; 1936; Ecological relations of ponerinae and other ants to termites; Proceedings of the American Academy of Arts and Sciences; 21 (14): 173-181 Wheeler, W. M. (1936). Ecological relations of ponerine and other ants to termites. Proceedings of the American Academy of Arts and Sciences, 71: 159-243
|
||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|