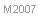|
|||||||||||||||||||||||||||||||||||||||||||||||||||
|
|||||||||||||||||||||||||||||||||||||||||||||||||||
| Género
Aphaenogaster Mayr, 1853
©Kiko Gómez & Xavier Espadaler |
Similares a Messor y Stenamma, se dintinguen de Stenamma ya que éste presenta ojos mucho menores y la separación entre lóbulos frontales menor que la anchura de la antena. La separación de Messor no está clara en las obreras, si bien analizando los machos no cabe error posible. Como ejemplo de la confusión entre estos dos géneros citar la especie A. striativentris, presente en la Península y que presenta una convergencia evolutiva muy clara con las especies del género Messor. En Bolton (1994) ambos géneros se separan en base a las mandíbulas (caracter no válido para el caso citado) y en la presencia o no de un proceso metaesternal. Una discusión acerca de como separar Messor y Aphaenogaster en la casta obrera se encuentra en Tinaut (1990). En ella se propone la longitud relativa del escapo respecto del ancho de la cabeza como indicador (LE/AC*100>128 para Aphaenogaster) en vez del que propone Bolton de Índice cefálico (ancho/longitud cabeza*100>90 para Messor). Sin embargo, en el caso de obreras aisladas ninguno de los dos métodos resulta efectivo al 100%. A pesar de su evidente engorro, hemos preferido a presencia o no del proceso metaesternal para la separación de ambos géneros en las obreras, si bien en la mayoría de los casos un análisis biométrico de la media de la muestra servirá para la separación de los mismos.
En los machos de Aphaenogaster, la volsella del aparato genital es muy reducida, prácticamente vestigial, mientras que en Messor volsella y lacinia presentan un tamaño similar (Tinaut, 1990).
En la Península Ibérica están representados dos subgéneros. Aphaenogaster (s. str.) y Attomyma.
Se han citado varias especies que consideramos debería ratificarse su presencia antes de incluirla de manera oficial en la Fauna Penínsular. Casi todas son citas del siglo XIX de especies que en la actualidad se consideran restingidas a la zona Mediterránea de Argelia y Túnez.
Se conocen más de 160 especies y más de una decena de especies fósiles distribuidas por todo el mundo excepto Sudamérica y África del Sur. En la Península Ibérica, si bien existen especies como A. subterranea que habitan preferentemente en la mitad Norte, las espeices de este género tienden a evitar la zona Norte y Noroeste de la Península Ibérica, aumentando el número de especies hacia el Sur y el Mediterráneo. Ciertas especies como A. striativentris o A. splendida solamente han sido descubiertas hasta el momento del tercio Sur peninsular. En cuanto a su origen, se pueden agrupar en:
Estas homigas nidifican siempre en el suelo. Sus nidos suelen estar provistos de un gran número de entradas en un espacio relativamente pequeño. Las entradas pueden estar o no cubiertas de piedrecillas u otros objetos, y en caso de estar excavado en arena o terreno blando, presentan forma de embudo que llega a tener 4 cm de diámetro y 30 cm de profundidad, rodeadas de volcanes de desperdicios. En Australia en ciertos lugares como campos de golf, pastos, etc, llegan a ser un serio problema, ya que socavan el terreno haciéndolo inestable. En terrenos duros las entradas son mucho menores, con pequeñas barreras de desperdicio rodeándola. Las del grupo Novomessor presentan diseños en forma de túmulos, similares a los nidos de Formica pero menores en tamaño, empleados para regular la temperatura (Holldobler & Wilson 1990). Los nidos no son únicos para cada colonia, sino que migran frecuentemente de una a otra localización. En el caso de A. senilis se ha comprobado que prácticamente todas las colonias obervadas en Doñana trasladan sus nidos de zonas insoladas en invierno a lugares más resguardados a la sombra en verano (Angel Barroso com. per.). En el caso de la norteamericana A. rudis se ha constado que la presencia en un nido varía entre 19 y 37 días (Smallwood 1982, Herbers 1985). En caso de ser molestadas saldrán unas pocas obreras a defender el nido. El número de individuos por nido es muy variable, siendo habitual una población del orden de varios cientos de obreras. Como en otros géneros, las obreras ponen huevos tróficos con los que alimentan a las larvas o la reina (Holldobler&Wilson 1990). Éstos suelen diferir de los de la reina en ser más flácidos y sin ADN detectable. Las larvas son alimentadas también con artrópodos. En el caso de que la presa sea demasiado grande, las hormigas optan por poner las larvas sobre la presa para que se alimenten de ella. Las reinas pueden vivir varios años-en cautividad, hasta 13 años para A. rudis (Haskins 1960). Estas hormigas no son agresivas, y lo compensan con un buen sistema de reclutamiento para que no le sean arrebatadas las presas. A. albisetosus y A. cockerelli, de los desiertos de Norteamérica, emplean la estridulación como un reforzador en la comunicación (Hölldobler et al. 1978, Markl & Hölldobler 1978). Así, para reforzar la señal de llamada hacia una presa estas hormigas, además de emitir las feromonas típicas de la glándula del veneno, estridulan para que las compañeras acudan antes. Se calcula que de este modo ganan entre uno y dos minutos a sus competidores. Esta segunda especie emplea un método más expeditivo para bloquear el forrajeo de su competidora Pogonomymex barbatus, ¡cerrándoles el nido!, con lo que les retrasan entre una y tres horas (Gordon 1988). También se ha demostrado para estas dos especies la presencia de feromonas de alarma que sirve para provocar la emigración del nido ante la presencia de las hormigas legionarias (Holldobler & Wilson 1990). Se ha demostrado también que la respuesta de las especies de este subgénero hacia las hormigas legionarias del género Neivamyrmex se ve incrementada tras un contacto continuado (McDonald & Topoff 1986) Las obreras de diferentes especies -A. senilis (Ledoux & Dargagnon 1973) y A. subterranea (Bruniquel 1972)- en nuestra fauna pueden llegar a reproducirse bien siendo inseminadas, bien telitokóticamente, esto es, generando obreras o reinas a partir de huevos no fecundados. Las reinas de diversas especies generan feromonas inhibidoras del desarrollo de los ovarios de las hembras, para que no puedan ejercer de competidoras de la reina, como se ha demostrado para A. cockerelli (Holldobler & Wilson 1990). Como parece ser la norma en muchos otros géneros, los rastros dejados por estas hormigas hacia la comida suelen ser específicos de cada nido, esto es, una hormiga reconoce si el rastro detectado es de hormigas de su nido o no (Hölldobler & Wilson 1990). Se alimentan tanto de semillas como de restos animales, si bien existen especies más especializadas en la recolección de semillas, como A. striativentris en la Peninsula Ibérica o el subgénero Novomessor en Norteamérica. Dentro de este género se encuentran las "hormigas jirafa" Aphaenogaster (Deromyrma) phalangium de centroamérica, con los cuellos inusualmente largos. Esta característica les permite navegar a través de charcos de agua, lo que les hace las primeras recolectoras tras las lluvias (Holldobler & Wilson 1990). Se ha descrito al menos una especie del Este de los EEUU, A. tennesseensis, inquilina temporal, que parasita a A. fulva (Creighton 1950).
En lo que incumbe a la fauna paleártica la mayoría de los trabajos son de comienzos del siglo XX, y desfasados en su mayor parte. Emery en 1908 propone una revisión del género para la fauna paleártica, que si bien obsoleta, merece (como la mayoría de sus trabajos) ser estudiada. En un trabajo posterior (Emery 1915) define los subgéneros, incluida la definición formal de Attomyrma (o grupo subterranea). Finzi (1930) realiza una revisión del subgénero Attomyrma que se ve complementado por el trabajo de Sanstchi (1933) para el subgénero Aphaenogaster (s. str.). No existe ningún trabajo reciente que revise el género para la fauna Mediterránea, aunque sí existen varios trabajos importantes sobre el género, en particular toda la serie de H. Cagniant sobre las Aphaenogaster norteafricanas con más de 20 artículos sobre esta fauna en Marruecos, Túnez y Argelia, que culminan con su revisión de las Aphaenogaster de Marruecos (1996), que si bien no incluye toda la fauna norteafricana sí es un artículo de apoyo importante para la comprensión de la fauna ibérica. Existen claves para la Europa mediterránea excluyendo la Península Ibérica realizadas por D. Agosti y C. A. Collingwood (1987) En otras zonas geográficas, Arnoldi (1976) revisa la fauna de la URRS, Park y Kim (2000) la fauna coreana y Longino (2004) analiza el complejo de A. phalangium. No existe una revisión mundial ni regional (que afecte a la península ibérica) reciente. Agosti, D.; Collingwod, C. A. (1987). A provisional list of the Balkan ants (Hym. Formicidae) with a key to the worker caste. II. Key to the worker caste, including the European species without the Iberian. Bulletin de la Societé entomologique Suisse, 60: 261-293 Arnol'di, K. V. (1976). Review of the genus Aphaenogaster (Hymenoptera, Formicidae) in the USSR. [in Russian]. Zoologicheski Zhurnal [=Zoological Journal, Moscow], 55: 1019-1026 Bolton, B. (1994). Identification guide to the ant genera of the world. Cambridge, Mass.:Harvard University Press: 222 pp Bruniquel, S (1972). La Ponte de la Fourmi Aphoenogaster subterranea. (Latr.) : oeufs reproducteurs-oeufs alimentaires. Comptes Rendus de l'Académie des Sciences, Paris Série D Sciences Naturelles, 275: 397-399 Cagniant, H. (1996). Les Aphaenogaster du Maroc (Hymenoptera: Formicidae): clé et catalogue des espèces. Annales de la Société Entomologique de France (N.S.), 32: 67-85 Creighton, W. S. (1950). The ants of North America. Bulletin of the Museum of Comparative Zoology at Harvard University, 104: 1-585 + 57lam. Emery, C. (1908). Beiträge zur Monographie der Formiciden des paläarktischen Faunengebietes. (Hym.) (Fortsetzung.) III. Die mit Aphaenogaster verwandte Gattungengruppe. Deutsche Entomologische Zeitschrift, 1908: 305-338 Emery, C. (1915). Definizione del genere Aphaenogaster e partizione di esso in sottogeneri. Parapheidole e Novomessor nn. gg. Rendiconti delle Sessioni della Reale Accademia delle Scienze dell'Istituto di Bologna Classe di Scienze Fisische, (n.s.)19: 67-75 Espadaler, X. (1981 ("1979")). Una nueva hormiga de la Península Ibérica. Miscellanea Zoologica, 5: 77-81 Finzi, B. (1930). Contributo allo studio degli Aphaenogaster paleartici (Formicidae-Myrmicinae). Bollettino della Società Entomologica Italiana, 62: 151-156 Gordon, D. M. (1988). Nest-plugging: interference competition in desert ants (Novomessor cockerelli and Pogonomyrmex barbatus). Oecologia (Berlin), 75: 114-118 Haskins, C. P. (1960). Note on the natural longevity of fertile females of Aphaenogaster picea. Journal of the New York Entomological Society, 68: 66-67 Herbers,J. M.; Adamowicz, S. C.;Helms, S. D. (1985). Seasonal changes in social organization of Aphaenogaster rudis (Hymenoptera: Formicidae). Sociobiology, 10: 1-15 Hölldobler, B.; Stanton, R. C.; Markl, H. (1978). Recruitment and food-retrieving behavior in Novomessor (Formicidae, Hymenoptera). I. Chemical signals. Behavioral Ecology and Sociobiology, 4: 163-181 Hölldobler, Bert; Wilson, E.O. (1990). The Ants. Belknap - Harvard: 732 pp Ledoux, A.; Dargagnon, D.(1973). La formation des castes chez la fourmi Aphaenogaster senilis Mayr. Comptes Rendus de l'Académie des Sciences, Paris Série D Sciences Naturelles, 276: 551-553 Longino, J. T. (2004). A revision of the Aphaenogaster phalangium complex (Hymenoptera: Formicidae: Myrmicinae). Zootaxa, 655: 1-12 McDonald, P.; Topoff, H. (1986). The development of defensive behavior against predation by army ants. Developmental Psychobiology, 19: 351-367 Markl, H.; Hölldobler, B. (1978). Recruitment and food-retrieving behavior in Novomessor (Formicidae, Hymenoptera). II. Vibration signals. Behavioral Ecology and Sociobiology, 4: 183-216 Park, S. J.; Kim, B. J. (2000). Sistematic study of Pheidolini in Korea. Korean Journal of Entomology, 30 (2): 107-114 Santschi, F. (1933). Étude sur le sous-genre Aphaenogaster Mayr. Revue Suisse de Zoologie, 40: 389-408 Smallwood, J. (1982). Nest relocations in ants. Insectes Sociaux, 29: 138-147 Tinaut, A.; Jiménez Rojas, J. (1991 ("1990")). Redescripción de Aphaenogaster striativentris Forel, 1895 y consideraciones sobre su polimorfismo (Hymenotera, Formicidae). EOS (Revista española de entomología), 66 (2): 117-126 Tinaut, A.; Pascual, R. (1986). Confirmación de la presencia de Aphaenogaster splendida (Roger, 1859) en la Península Ibérica. Nouvelle Revue d'Entomologie, (n.s.)3 (2): 189-192
|
||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|